Hormony mózgowe pszczoły
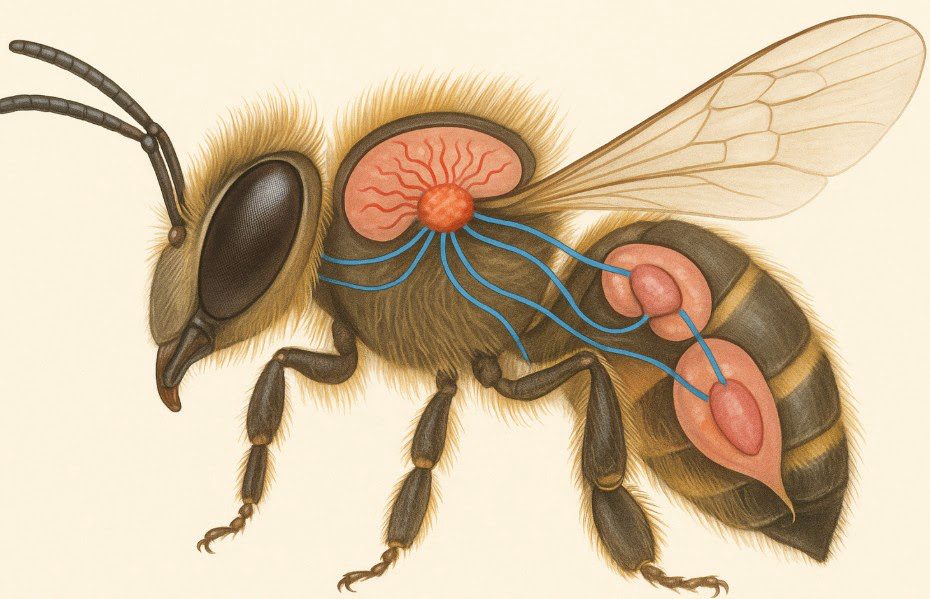
Hormony mózgowe pszczoły stanowią kluczowy element systemu neuroendokrynnego, odpowiadający za regulację złożonych procesów behawioralnych, uczenia się oraz koordynację funkcji społecznych w kolonii. Mózg pszczoły produkuje różnorodne neurohormony i neuroprzekaźniki, które modulują aktywność układu nerwowego oraz wpływają na inne gruczoły endokrynne w organizmie. System neurohormonalny integruje sygnały środowiskowe z wewnętrznymi potrzebami organizmu, umożliwiając precyzyjne dostosowywanie zachowań do zmieniających się warunków.
Złożoność funkcjonalna hormonów mózgowych odzwierciedla zaawansowaną organizację społeczną pszczół, gdzie konieczna jest precyzyjna koordynacja zachowań tysięcy osobników. Plastyczność neuralna regulowana przez hormony mózgowe umożliwia pszczołom uczenie się, zapamiętywanie tras zbierania oraz adaptację do nowych wyzwań środowiskowych. Komunikacja neurohormonalna odbywa się na wielu poziomach – od lokalnej modulacji synaps po systemową regulację zachowań całego organizmu.
Komórki neurosekrecyjne mózgu
Komórki neurosekrecyjne w mózgu pszczoły to wyspecjalizowane neurony zdolne do produkcji i wydzielania hormonów bezpośrednio do hemolimfy lub do innych części układu nerwowego. Część międzymózgowa w protocerebrum zawiera największe skupisko tych komórek, produkujących kluczowe neurohormony kontrolujące rozwój i zachowanie. Morfologia komórek neurosekrecyjnych różni się od zwykłych neuronów – posiadają one rozbudowane ciało komórkowe z licznymi organellami syntetyzującymi białka oraz długie aksony zakończone rozszerzonymi zakończeniami uwalniającymi hormony.
Rozmieszczenie komórek neurosekrecyjnych w różnych obszarach mózgu odzwierciedla ich specjalizację funkcjonalną – niektóre kontrolują gruczoły endokrynne, inne modulują zachowania społeczne lub procesy poznawcze. Środkowe komórki neurosekrecyjne produkują hormon prothoracicotropowy (PTTH) regulujący metamorfozę, podczas gdy boczne komórki neurosekrecyjne wydzielają neuropeptydy wpływające na zachowania i metabolizm. Cykliczna aktywność tych komórek jest synchronizowana z rytmami dobowymi oraz sezonowymi zmianami w koloni.
Ultrastruktura komórek neurosekrecyjnych ujawnia liczne ziarnistości wydzielnicze zawierające gotowe do uwolnienia hormony oraz rozbudowany aparat Golgiego i siateczkę endoplazmatyczną odpowiedzialne za syntezę i pakowanie neuropeptydów. Transport aksonalny przenosi hormony z miejsc syntezy do zakończeń uwalniających, gdzie są magazynowane w pęcherzykach wydzielniczych. Regulacja uwalniania hormonów odbywa się poprzez sygnały elektryczne oraz modulację przez inne neuroprzekaźniki i hormony.
Dopamina i kontrola zachowań społecznych
Dopamina w mózgu pszczoły pełni fundamentalną rolę w regulacji motywacji, uczenia się oraz kontroli zachowań społecznych, działając jako kluczowy neuroprzekaźnik w szlakach nagradzania i podejmowania decyzji. Neurony dopaminowe są rozmieszczone głównie w protocerebrum i unerwią obszary mózgu odpowiedzialne za przetwarzanie informacji sensorycznych i planowanie zachowań. Stężenie dopaminy w mózgu pszczoły wynosi około 1-5 nmol/g tkanki mózgowej, co czyni ją jednym z najważniejszych neuroprzekaźników w systemie nerwowym.
Feromon matki wywiera głęboki wpływ na szlaki dopaminowe w mózgu młodych robotnic, modulując ich poziom aktywności oraz skłonność do wykonywania określonych zadań w ulu. Kwas homowaniliowy (HVA), metabolit dopaminy, służy jako biomarker aktywności dopaminergicznej i jego stężenie zmienia się w odpowiedzi na sygnały feromonowe od matki. Receptory dopaminowe należące do rodziny receptorów sprzężonych z białkami G regulują aktywność drugich przekaźników takich jak cAMP i wapń wewnątrzkomórkowy.
Funkcja dopaminy w procesach uczenia się jest szczególnie wyraźna podczas formowania skojarzeń między zapachami kwiatów a nagrodami pokarmowymi. Modulacja plastyczności synaptycznej przez dopaminę umożliwia konsolidację wspomnień związanych z efektywnym zbieraniem pokarmu. Zaburzenia w systemie dopaminowym mogą prowadzić do problemów z orientacją, zmniejszonej motywacji do pracy oraz zaburzeń w hierarchii społecznej kolonii.
Oktopamina jako hormon stresu
Oktopamina funkcjonuje jako główny hormon stresu u pszczół, odpowiadając za mobilizację organizmu w sytuacjach zagrożenia oraz zwiększenie gotowości do aktywności fizycznej i obrony kolonii. Neurony oktopaminowe rozmieszczone w mózgu i zwojach obwodowych unerwią mięśnie, gruczoły i ośrodki nerwowe odpowiedzialne za reakcje stresowe. Mechanizm działania oktopaminy przypomina funkcje adrenaliny u kręgowców, aktywując procesy metaboliczne i zwiększając wrażliwość układu nerwowego na bodźce zewnętrzne.
Uwalnianie oktopaminy następuje w odpowiedzi na sygnały alarmowe, zagrożenia dla kolonii, aktywność zbierania pokarmu oraz podczas intensywnej pracy fizycznej. Receptory oktopaminowe modulują aktywność mięśni lotu, zwiększając ich wydajność podczas długich lotów zbierania oraz szybkość reakcji podczas ucieczki od drapieżników. Wpływ na układ nerwowy obejmuje zwiększenie przewodnictwa synaptycznego, wzmocnienie procesów uczenia się oraz poprawę zdolności koncentracji na zadaniach wymagających precyzji.
Wzajemne oddziaływanie oktopaminy z innymi neuroprzekaźnikami tworzy złożone sieci regulacyjne kontrolujące stan pobudzenia organizmu. Antagonistyczne działanie względem serotoniny zapewnia równowagę między stanami aktywacji i relaksacji. Modulacja produkcji oktopaminy przez czynniki środowiskowe takie jak temperatura, dostępność pokarmu czy obecność zagrożeń pozwala na adaptacyjne dostosowywanie odpowiedzi behawioralnych.
Serotonina i regulacja nastroju
Serotonina w mózgu pszczoły reguluje nastrój, agresywność oraz ogólny stan behawioralny, działając często antagonistycznie do oktopaminy w kontroli poziomu pobudzenia organizmu. Około 75 neuronów serotoninowych rozmieszczonych w płatach optycznych, środkowym protocerebrum oraz grzbietowym protocerebrum unerwiają większość obszarów mózgu z wyjątkiem kielichów ciałek grzybowatych. Włókna serotoninowe tworzą gęstą sieć w płatach optycznych, ciałkach zapachowych oraz centrum mózgu, modulując przetwarzanie informacji sensorycznych.
Funkcja serotoniny w regulacji cyklów aktywności jest szczególnie ważna dla synchronizacji zachowań pszczół z rytmami dobowymi i sezonowymi. Poziom serotoniny wzrasta podczas okresów odpoczynku i obniża się podczas intensywnej aktywności, przeciwdziałając efektom oktopaminy. Modulacja progów reakcji na bodźce społeczne przez serotoninę wpływa na podział pracy w kolonii oraz na skłonność do wykonywania określonych zadań.
Receptory serotoninowe należące do różnych podtypów regulują odmienne aspekty fizjologii i zachowania – receptory 5-HT1 hamują aktywność neuronalną, podczas gdy 5-HT2 zwiększają pobudliwość niektórych obszarów mózgu. Wzajemne oddziaływanie z systemem dopaminowym tworzy subtelne równowagi emocjonalne umożliwiające precyzyjną kontrolę zachowań społecznych. Zaburzenia w funkcjonowaniu serotoniny mogą prowadzić do zwiększonej agresywności, problemów z komunikacją społeczną oraz zaburzeń rytmów aktywności.
Tyramina i modulacja zachowań
Tyramina jest aminą biogenną o mniej poznanym mechanizmie działania, ale sugeruje się jej antagonistyczne funkcje względem oktopaminy w regulacji zachowań i procesów fizjologicznych u pszczół. Stężenie tyraminy w mózgu jest relatywnie niskie w porównaniu do innych bioamin, але jej obecność w kluczowych obszarach neuropilu wskazuje na ważną rolę modulacyjną. Receptory tyraminy należą do rodziny receptorów sprzężonych z białkami G i wpływają na poziomy drugich przekaźników wewnátrzkomórkowych.
Funkcja hamująca tyraminy może przeciwdziałać nadmiernej stymulacji przez oktopaminę, zapewniając mechanizmy ujemnego sprzężenia zwrotnego w kontroli poziomu pobudzenia. Wpływ na plastyczność synaptyczną może modulować zdolności uczenia się oraz utrwalanie pamięci długotrwałej. Ekspresja genów kodujących enzymy syntezy tyraminy zmienia się w zależności od wieku pszczoły i wykonywanych funkcji społecznych.
Badania farmakologiczne z użyciem agonistów i antagonistów tyraminy ujawniają jej wpływ na motywację do pracy, wrażliwość na nagrody pokarmowe oraz skłonność do eksploracji nowych środowisk. Wzajemne oddziaływania z innymi systemami neuroprzekaźników tworzą złożone sieci regulacyjne umożliwiające subtelną modulację zachowań. Rola w komunikacji społecznej może obejmować wpływ na wrażliwość na feromony i inne sygnały chemiczne.
Histamina w systemie wzrokowym
Histamina w mózgu pszczoły jest głównym neuroprzekaźnikiem w systemie wzrokowym, z około 150 neuronami histaminowymi unerwiającymi większość obszarów protocerebrum z wyjątkiem ciałek grzybowatych. Fotoreceptory w oczach złożonych oraz oczka proste wykorzystują histaminę do przekazywania sygnałów świetlnych do wyższych ośrodków przetwarzania wzrokowego. Włókna histaminowe zakończają się w blaszce i rdzeniu płatów optycznych, gdzie modulują przetwarzanie informacji o ruchu, kolorach i wzorcach.
Mechanizm synaptyczny histaminy opiera się na aktywacji receptorów chlorkowych, które powodują hiperpolaryzację błon postsynaptycznych i tym samym hamowanie aktywności neuronalnej. Szybka kinetyka przekaźnictwa histaminowego umożliwia precyzyjne czasowe kodowanie sygnałów wzrokowych niezbędne dla analizy ruchu i kontroli lotu. Plastyczność synaps histaminowych może uczestniczyć w adaptacji do różnych warunków świetlnych oraz w uczeniu się wzorców wzrokowych.
Farmakologiczna manipulacja systemu histaminowego wpływa na zdolności orientacji przestrzennej, rozpoznawanie punktów orientacyjnych oraz efektywność zbierania pokarmu. Wzajemne oddziaływanie z innymi systemami neuroprzekaźników w płatach optycznych moduluje wrażliwość na różne aspekty bodźców wzrokowych. Rozwój ontogenetyczny systemu histaminowego współgra z dojrzewaniem funkcji wzrokowych podczas metamorfozy.
Neuropeptydy regulacyjne
Neuropeptydy w mózgu pszczoły stanowią największą grupę sygnalizatorów chemicznych, obejmującą dziesiątki różnych związków o różnorodnych funkcjach regulacyjnych w kontroli zachowań, metabolizmu i rozwoju. Peptydy insulinopodobne (ILP) regulują wzrost, metabolizm oraz długość życia, działając podobnie do insuliny u kręgowców w kontroli homeostazy glukozowej. Neuropeptyd F (NPF) wpływa na apetyt, motywację do poszukiwania pokarmu oraz zachowania społeczne związane z podziałem pracy.
Allatostatyna i allatotropina regulują aktywność ciałek przyległych oraz produkcję hormonu juwenilnego, stanowiąc kluczowe elementy kontroli endokrynnej rozwoju i reprodukcji. Corazonina wpływa na metabolizm energetyczny, mobilizację zapasów lipidowych oraz zachowania związane z aktywnością lokomocyjną. Tachykininy modulują aktywność mięśni gładkich w przewodzie pokarmowym oraz wpływają na zachowania eksploracyjne.
PBAN (neuropeptyd aktywujący biosyntezę feromonów) kontroluje produkcję feromonów w gruczołach wydzielniczych, regulując komunikację chemiczną w kolonii. Krótki neuropeptyd F (sNPF) moduluje apetyt, zachowania żywieniowe oraz reakcje na stres. Badania funkcjonalne neuropeptydów wykorzystują techniki RNAi, immunohistochemię oraz analizy behawioralne po aplikacji syntetycznych peptydów.
Acetylocholina w procesach poznawczych
Acetylocholina w mózgu pszczoły pełni kluczową rolę w procesach poznawczych, uczeniu się oraz formowaniu pamięci, szczególnie w ciałkach grzybowatych odpowiedzialnych za skomplikowane formy uczenia skojarzeniowego. Neurony cholinergiczne unerwią większość obszarów mózgu, z najgęstszą unerwacją w kielichach ciałek grzybowatych gdzie zachodzą procesy integracji wielomodalnej. Receptory nikotynowe i muskarynowe acetylocholiny regulują różne aspekty plastyczności synaptycznej oraz procesów pamięciowych.
Funkcja acetylocholiny w uczeniu się skojarzeń zapach-nagroda jest szczególnie dobrze udokumentowana w badaniach elektrofizjologicznych i behawioralnych. Modulacja aktywności neuronów Kenyon w ciałkach grzybowatych przez acetylocholinę umożliwia formowanie specyficznych reprezentacji zapachowych oraz ich kojarzenie z wartością motywacyjną. Plastyczność synaptyczna zależna od acetylocholiny pozwala na długotrwałe zmiany w siłach połączeń neuronalnych.
Degradacja acetylocholiny przez acetylocholinoesterazę kontroluje czasowy przebieg sygnalizacji cholinergicznej, umożliwiając precyzyjną kontrolę procesów poznawczych. Inhibitory acetylocholinoesterazy wpływają na zdolności uczenia się i pamięć, co ma znaczenie w kontekście toksycznego działania niektórych pestycydów. Rozwój ontogenetyczny systemu cholinergicznego współgra z dojrzewaniem zdolności poznawczych podczas metamorfozy.
GABA i hamowanie neuralne
GABA (kwas gamma-aminomasłowy) stanowi główny neuroprzekaźnik hamujący w układzie nerwowym pszczoły, regulując pobudliwość neuronów oraz zapobiegając nadmiernej aktywacji obwodów neuronalnych. Neurony GABAergiczne są rozmieszczone w większości obszarów mózgu, szczególnie gęsto w płatach optycznych, ciałkach zapachowych oraz obszarach motorycznych. Receptory GABA należą głównie do typu jonotropowego sprzężonego z kanałami chlorkowymi, powodując hiperpolaryzację błon neuronalnych.
Funkcja regulacyjna GABA w kontroli rytmów neuronalnych jest kluczowa dla prawidłowego przetwarzania informacji sensorycznych oraz koordynacji ruchów. Równowaga między pobudzeniem cholinergicznym a hamowaniem GABAergicznym determinuje wrażliwość obwodów neuronalnych na bodźce zewnętrzne. Modulacja siły hamowania GABAergicznego wpływa na progi reakcji oraz selektywność odpowiedzi behawioralnych.
Zaburzenia w funkcjonowaniu systemu GABAergicznego mogą prowadzić do napadów drgawkowych, nieprawidłowych wzorców aktywności ruchowej oraz problemów z kontrolą zachowań impulsywnych. Farmakologiczna manipulacja receptorów GABA wpływa na zdolności uczenia się, poziom lęku oraz ogólną aktywność behawioralną. Wzajemne oddziaływania z innymi systemami neuroprzekaźników tworzą złożone sieci regulacyjne kontrolujące homeostazę neuronalną.
Glutamian jako główny neuroprzekaźnik pobudzający
Glutamian jest najważniejszym neuroprzekaźnikiem pobudzającym w układzie nerwowym pszczoły, odpowiedzialnym za szybką transmisję synaptyczną w większości połączeń neuronalnych. Receptory glutaminanu typu AMPA i NMDA regulują podstawowe procesy przekaźnictwa oraz plastyczność synaptyczną niezbędną dla uczenia się i pamięci. Gęstość receptorów glutaminanu jest szczególnie wysoka w obszarach odpowiedzialnych za przetwarzanie informacji sensorycznych oraz kontrolę motoryczną.
Mechanizmy plastyczności zależnej od glutaminanu obejmują długotrwałe wzmocnienie (LTP) i osłabienie (LTD) synaptyczne, które są podstawą molekularną procesów uczenia się i zapamiętywania. Aktywacja receptorów NMDA wymaga jednoczesnej depolaryzacji błony postsynaptycznej oraz obecności glutaminanu, co czyni je idealnymi detektorami zbieżności potrzebnej do formowania skojarzeń. Homeostaza glutaminanu jest regulowana przez transportery wychwytu zwrotnego oraz enzymy metabolizujące.
Nadmierna aktywacja receptorów glutaminanu może prowadzić do ekscytotoksyczności – uszkodzenia neuronów przez nadmierny napływ wapnia i aktywację procesów apoptotycznych. Modulacja funkcji glutaminanu przez inne neuroprzekaźniki pozwala na subtelną regulację siły synaptycznej w zależności od kontekstu behawioralnego. Rozwój systemu glutaminanu podczas metamorfozy współgra z dojrzewaniem funkcji poznawczych i motorycznych.
Wzajemne oddziaływania między systemami neuroprzekaźnikowymi
Złożone wzajemne oddziaływania między różnymi systemami neuroprzekaźników w mózgu pszczoły tworzą dynamiczne sieci regulacyjne umożliwiające precyzyjną kontrolę zachowań i procesów poznawczych. Komunikacja krzyżowa między systemami dopaminowym i serotoninowym reguluje równowagę między motywacją a hamowaniem behawioralnym. Modulacja aktywności cholinergicznej przez aminy biogenne wpływa na efektywność procesów uczenia się w różnych kontekstach motywacyjnych.
Współmodulacja przez neuropeptydy wprowadza dodatkowy poziom regulacji, umożliwiając długotrwałe zmiany w responsywności obwodów neuronalnych. Plastyczność na poziomie receptorów pozwala na adaptacyjne zmiany w wrażlivości na różne neuroprzekaźniki w odpowiedzi na doświadczenie. Rytmiczne zmiany w poziomach różnych neuroprzekaźników synchronizują procesy behawioralne z cyklami dobowymi i sezonowymi.
Farmakologiczne interwencje celujące w liczne systemy neuroprzekaźników ujawniają właściwości emergentne sieci neuronalnych niemożliwe do przewidzenia na podstawie analizy pojedynczych komponentów. Modelowanie obliczeniowe wzajemnych oddziaływań neuroprzekaźników może pomóc w zrozumieniu zasad organizacji i funkcjonowania mózgu pszczoły. Pochodzenie ewolucyjne różnych systemów neuroprzekaźników wskazują na ich fundamentalne znaczenie w kontroli zachowań społecznych.
Wpływ czynników środowiskowych na hormony mózgowe
Temperatura ma bezpośredni wpływ na aktywność enzymów syntetyzujących i degradujących neuroprzekaźniki, wpływając na ich stężenia i efektywność działania w mózgu. Stres termiczny może zakłócać równowagę między różnymi systemami neuroprzekaźników, prowadząc do zaburzeń behawioralnych i poznawczych. Optymalna temperatura około 35°C w obszarze czerwiu jest kluczowa dla prawidłowego rozwoju systemu nerwowego i produkcji hormonów mózgowych.
Dostępność pokarmu wpływa na produkcję hormonów mózgowych poprzez modulację stanu metabolicznego oraz dostępność prekursorów aminokwasowych niezbędnych do syntezy neuroprzekaźników. Okresy głodu prowadzą do zmian w profilach neuroprzekaźników, przesuwając priorytety behawioralne w kierunku poszukiwania pokarmu. Jakość pyłku i zawartość białek w diecie wpływają na syntezę neuropeptydów oraz ogólną homeostazę neuroprzekaźnikową.
Zanieczyszczenia środowiskowe mogą działać jako neurotoksyny, zakłócając normalne funkcjonowanie systemów neuroprzekaźników i prowadząc do zaburzeń behawioralnych. Pestycydy szczególnie te z grupy neonikotynoidów, mogą interferować z receptorami acetylocholiny, prowadząc do problemów z uczeniem się i pamięcią. Metale ciężkie mogą zakłócać funkcjonowanie enzymów niezbędnych do syntezy i metabolizmu neuroprzekaźników.
Technologie badawcze hormonów mózgowych
Wysokosprawna chromatografia cieczowa sprzężona ze spektrometrią mas (HPLC-MS) stanowi złoty standard w analizie neuroprzekaźników i neuropeptydów w próbkach mózgowych. Mikrodializa in vivo umożliwia monitorowanie dynamicznych zmian stężeń neuroprzekaźników w żywych mózgach pszczół podczas różnych aktywności behawioralnych. Czujniki elektrochemiczne pozwalają na pomiary w czasie rzeczywistym niektórych neuroprzekaźników z wysoką rozdzielczością czasową.
Immunohistochemia z użyciem specyficznych przeciwciał umożliwia lokalizację określonych neuroprzekaźników i ich receptorów w strukturach mózgowych. Hybrydyzacja in situ pozwala na mapowanie ekspresji genów kodujących enzymy syntezy neuroprzekaźników oraz ich receptory. Podejścia transgeniczne z użyciem fluorescencyjnych reporterów umożliwiają śledzenie aktywności określonych populacji neuronów in vivo.
Interwencje farmakologiczne z użyciem agonistów, antagonistów oraz inhibitorów enzymatycznych pozwalają na charakterystykę funkcjonalną różnych systemów neuroprzekaźników. Testy behawioralne łączą manipulacje farmakologiczne z ilościową analizą zachowań, ujawniając funkcjonalne znaczenie specyficznych szlaków neuronalnych. Rejestracje elektrofizjologiczne bezpośrednio mierzą wpływ neuroprzekaźników na aktywność neuronalną in vitro i in vivo.
Znaczenie dla pszczelarstwa i ochrony pszczół
Zrozumienie hormonów mózgowych ma kluczowe znaczenie dla opracowywania strategii ochrony pszczół oraz monitorowania ich zdrowia neurobiologicznego. Biomarkery oparte na profilach neuroprzekaźników mogą służyć jako wczesne wskaźniki narażenia na neurotoksyny lub stres środowiskowy. Analiza zachowań związanych ze specyficznymi systemami neuroprzekaźników może pomóc w diagnozowaniu problemów zdrowotnych kolonii.
Monitorowanie poziomów hormonów związanych ze stresem jak oktopamina może wskazywać na problemy w zarządzaniu pasieką lub czynniki stresowe środowiskowe wpływające na dobrostan pszczół. Testy behawioralne badające funkcje uczenia i pamięci mogą wykrywać podletalne efekty pestycydów zanim pojawią się widoczne objawy. Zrozumienie rytmów dobowych hormonów mózgowych może zoptymalizować timing praktyk zarządzania.
Programy edukacyjne dla pszczelarzy na temat neurobiologii pszczół mogą przyczynić się do lepszego zrozumienia zachowań pszczół i poprawy zarządzania ulem. Programy hodowli selektywnej mogą uwzględniać cechy neurobiologiczne związane z odpornością na stres i zdolnościami poznawczymi. Opracowanie protokołów testowania neurotoksyczności może pomóc w ocenie bezpieczeństwa nowych pestycydów i innych chemikaliów.
FAQ
Jakie są najważniejsze neuroprzekaźniki w mózgu pszczoły?
Najważniejsze to acetylocholina (uczenie się i pamięć), dopamina (motywacja i nagradzanie), oktopamina (stres i pobudzenie), serotonina (nastrój i cykle aktywności), GABA (hamowanie) oraz glutamian (podstawowa transmisja pobudzająca).
Jak pestycydy wpływają na hormony mózgowe pszczół?
Pestycydy, szczególnie neonikotynnoidy, mogą blokować receptory acetylocholiny, zaburzając procesy uczenia się i pamięci. Mogą również wpływać na inne systemy neuroprzekaźników, prowadząc do problemów behawioralnych.
Czy można mierzyć poziomy hormonów mózgowych u żywych pszczół?
Tak, używając technik mikrodializy in vivo, czujników elektrochemicznych czy analiz hemolimfy. Jednak większość precyzyjnych analiz wymaga pobrania próbek tkanki mózgowej.
Jak wiek pszczoły wpływa na hormony mózgowe?
Profile neuroprzekaźników zmieniają się z wiekiem – młode pielęgniarki mają inne poziomy dopaminy i oktopaminy niż starsze zbieraczki, co współgra z ich różnymi funkcjami społecznymi.
Czy stres wpływa na hormony mózgowe kolonii?
Tak, stres kolonii prowadzi do wzrostu poziomów oktopaminy (hormonu stresu) oraz zmian w innych systemach neuroprzekaźników, wpływając na zachowania obronne i mobilizację pszczół.
Jak badać wpływ hormonów mózgowych na zachowanie pszczół?
Poprzez eksperymenty farmakologiczne (podawanie agonistów/antagonistów), analizy behawioralne po manipulacjach chemicznych oraz korelacje między poziomami neuroprzekaźników a obserwowanymi zachowaniami.
Czy można wpływać na hormony mózgowe pszczół w celach praktycznych?
W ograniczonym zakresie tak, ale wymaga to ostrożności ze względu na złożoność systemu i możliwe skutki uboczne. Bardziej praktyczne jest zarządzanie warunkami środowiskowymi wpływającymi na te hormony.
Jak temperatura wpływa na działanie hormonów mózgowych?
Temperatura bezpośrednio wpływa na aktywność enzymów syntetyzujących neuroprzekaźniki – zbyt niska lub wysoka może zaburzać ich produkcję i metabolizm, wpływając na zachowania i funkcje poznawcze.
Dodaj komentarz